



Bacillariophytina
David G. Mann



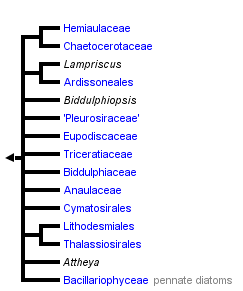


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Bacillariophytina is a subphylum (subdivision) of diatoms distinguished by Medlin & Kaczmarska (2004) on the basis of (1) molecular phylogenies derived from nuclear and plastid SSU rDNA and (2) aspects of cell shape, reflecting how the auxospore develops. It includes some of the diatoms with a centric valve structure and all diatoms with a pennate valve structure.
It is all but certain from molecular phylogenies that the centric diatoms are not a monophyletic group. Molecular data indicate that the monophyletic Bacillariophyceae (pennate diatoms) are not the sister group to all of the centric diatoms but evolved in one clade out of the several or many major clades of centric diatoms, and fossil evidence tends to confirm this. Although the exact relationships among the various clades of centrics, and between any of these and the pennate diatoms (= Bacillariophyceae), are currently unclear, many molecular phylogenies suggest the existence of a monophyletic group comprising the pennate diatoms and a particular subset of the centric diatoms that can be referred to as the 'polar centrics'. Together, these comprise the Bacillariophytina.
The Bacillariophytina (with one major exception) are characterized by having bi- or multipolar valves. Usually, this polarity is expressed in the shape and structure of the valves, but sometimes it is expressed in the structure alone. Thus, unlike the 'radial centric diatoms', which almost always have circular valves, the polar centrics and pennate diatoms are elliptical, elongate, lemon-like, banana-like, triangular, square, stellate, etc. In the polar centrics and pennates that do have circular or almost circular valves, these usually have obvious bi- or multipolar pattern, such as the possession of two or three or four special submarginal areas of secretory pores, often raised on stubby projections.
The capacity to produce non-circular valves seems usually to be associated with a special type of auxospore wall containing either a 'properizonium' (polar centric diatoms) or a 'perizonium' (pennate diatoms), which are complex systems of silicified strips laid down sequentially as the auxospore expands. (However, there are exceptions, scattered through the pennate diatoms, in which anisometric expansion occurs in auxospores with organic walls, apparently lacking perizonia). The properizonium and perizonium were first studied in detail by von Stosch (1962, 1982) and Mann (1982) and the correlation of auxospore coverings with phylogeny has been explored by Kaczmarska et al. (2001) and Medlin & Kaczmarska (2004).
The principal exception to bi- or multipolarity in the Bacillariophytina is the Thalassiosirales, a large group of centric diatoms that usually lack bi- or multi-polar pattern and usually have circular valves. Most molecular phylogenies published so far show the Thalassiosirales among the polar centric diatoms and the tree topologies (often with the Lithodesmiales as the sister group) make it most likely that the Thalassiosirales have lost bi- or multipolarity, rather than never having had it. It may be significant that a few Thalassiosirales, including Cylcotella iris (slightly elliptical) and C. austriaca (linear elliptical), have noncircular valves (Hustedt 1927-66, Serieyssol 1984) and these species may perhaps have retained a plesiomorphic type of auxospore wall containing a properizonium. However, molecular phylogenies (e.g. Alverson et al. 2007) do not support an early branching of Cyclotella from other Thalassiosirales. So, if Cyclotella is monophyletic and if the non-circular valves of C. iris and C. austriaca do reflect the presence of a functional properizonium, then either the circular-valved ancestors of these species retained a non-functional properizonium that was later restored to 'working order' to create elliptical outlines, or there must have been multiple losses of the properizonium in other Thalassiosirales clades.
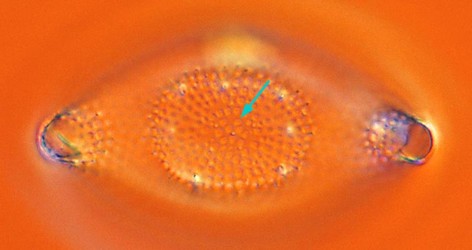
Odontella. © 2008 David G. Mann. Despite the elongate or multipolar outline of many polar centrics, the pattern-centre is usually a simple annulus (arrow) and the ribs and striae of the valve radiate out from this, as in radial centrics. The annulus is generally circular, as here, or elliptical, but is elongate or dumbbell-shaped in a few species, which may hint at how the sternum of pennate diatoms may have evolved. In the extremely elongate cells of the Ardissoneales, the pattern-centre is more complex.
The polar centric diatoms were given formal status by Medlin & Kaczmarska (2004), as the class Mediophyceae, alongside the class Bacillariophyceae (pennate diatoms). These two classes were also listed by Adl et al. (2005), with a clear indication that the Mediophyceae are most likely paraphyletic.
Characteristics
The only unifying feature of the Bacillariophytina separating them from the radial centric diatoms, apart from molecular synapomorphies, is the ability of these organisms to produce bi- or multipolar cells.
It is possible that no diatom valve is without some kind of polarity. Thus, even the radial centrics often have singular structures, e.g. the single submarginal pseudonodulus of Actinocyclus, the single narrow ray of Asteromphalus, or the slightly offset terminal processes of Rhizosolenia and Proboscia, which define an axis of symmetry. It might be more appropriate to call these diatoms 'monopolar'. The monopolar axis may perhaps bear a constant relationship to the position of the mitotic spindle and nucleus, which, as far as is known, always lie just beneath the girdle on one side of the cell, but this has not been investigated.
In contrast to monopolar/radial centric diatoms, most polar centrics and pennate diatoms have an obvious bi-, tri- or quadripolar (rarely >4) organization. The characteristics of the polar centric clades (= Medlin & Kaczmarska's Mediophyceae, comprising all the groups on the branches of the tree shown above, apart from the Bacillariophyceae) are:
- cells chain-forming or (less commonly) solitary
- valves usually bi- or multipolar in their structure, outline, or both
- distance between valves (pervalvar depth of the girdle) usually short
- valve pattern subtended by an annulus (sometimes highly modified)
- discrete submarginal pore fields (areas of porelli = unoccluded pores) often present, involved in secretion of polysaccharide stalks or pads; fultoportulae and chitin secretion present in Thalassiosirales
- rimoportulae usually present
- auxospore wall containing a properizonium, at least in those species producing non-circular valves (but the number of genera that have been investigated is small)
- planktonic or benthic
The characteristics of the Bacillariophyceae are similar except that:
- chain formation is less common
- the valves are almost always bipolar: multipolar forms are rare, occurring sporadically in various unrelated lineages
- the valve pattern is subtended by a sternum, not an annulus
- rimoportulae are present in some clades; however, in a major subclade (containing more species than all other centric and pennate clades) the principal wall organelles are not rimoportulae but a system of slits and associated cytoplasmic structures involved in locomotion, called the raphe system
- the auxospore wall contains a perizonium (except where this has been lost secondarily)
Fossil History
Polar centric diatoms are present in the first well-preserved, diverse diatom assemblage known at present – an Early Cretaceous marine deposit in the Weddell Sea, Antarctica (Gersonde & Harwood 1990, Sims et al. 2006). Earlier deposits only contain radial centrics, but the total number of taxa in these deposits is very low, so that the absence of polar centrics might be fortuitous. At present, therefore, the fossil record does not provide unambiguous evidence concerning the symmetry and shape of the ancestral diatoms. Semina & Beklemishev (1981) suggested on quasi-theoretical grounds that the radial centric construction, especially the 'indefinite polyaxonic' condition ("there is a centre of symmetry and a large but finite number of axes of symmetry") is primitive. However, If Medlin is correct in the claim that the extant radial centric diatoms comprise a monophyletic grouping sister to the polar centrics , pennates (e.g. Medlin & Kaczmarska 2004, p. 249; fig. 2 in Sims et al. 2006), then the absence of fossil evidence makes it virtually impossible to reconstruct the shape and symmetry of the ancestral diatoms, because the immediate relatives of diatoms (bolidophytes and possibly parmophytes) do not possess similar cell wall construction.
Pennate diatoms are known from the Late Cretaceous.
References
Alverson, A.J., Jansen, R.K. & Theriot, E.C. (2007). Bridging the Rubicon: phylogenetic analysis reveals repeated colonizations of marine and fresh waters by thalassiosiroid diatoms. Molecular Phylogenetics and Evolution 45: 193-210.
Gersonde, R. & Harwood, D.M. (1990). Lower Cretaceous diatoms from ODP Leg 113 site 693 (Weddell Sea) Part 1: vegetative cells. Proceedings of the Ocean Drilling Program, Scientific Results 113: 365402.
Hustedt, F. (192766). Die Kieselalgen Deutschlands, Österreichs und der Schweiz unter Berücksichtigung der übrigen Länder Europas sowie der angrenzenden Meeresgebiete. In Dr. L. Rabenhorsts Kryptogamenflora von Deutschland, Österreich und der Schweiz, vol. 7. Akademische Verlagsgesellschaft, Leipzig.
Kaczmarska, I., Ehrman, J.M. & Bates, S.S. (2001). A review of auxospore structure, ontogeny and diatom phylogeny. In: Proceedings of the 16th International Diatom Symposium (Ed. by A. Economou-Amilli), pp. 153-168. Amvrosiou Press, Athens.
Mann, D.G. (1982). Structure, life history and systematics of Rhoicosphenia (Bacillariophyta) II. Auxospore formation and perizonium structure of Rh. curvata. Journal of Phycology 18: 264-274.
Medlin, L.K. & Kaczmarska, I. (2004). Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245270.
Round, F.E., Crawford, R.M. & Mann, D.G. (1990). The diatoms. Biology and morphology of the genera. Cambridge University Press, Cambridge. 747 pp.
Semina, H.J. & Beklemishev, C.W. (1981). A promorphological approach to diatom cell structure. In: Proceedings of the 6th Symposium on Recent and Fossil Diatoms (Ed. by R. Ross), pp. 211-230. Otto Koeltz, Koenigstein.
Sims, P.A., Mann, D.G. & Medlin, L.K. (2006). Evolution of the diatoms: insights from fossil, biological and molecular data. Phycologia 45: 361-402.
Sorhannus, U. (2007). A nuclear-encoded small-subunit ribosomal RNA timescale for diatom evolution. Marine Micropaleontology 65: 1-12.
Stosch, H.A. von (1962). Über das Perizonium der Diatomeen. Vorträge aus dem Gesamtgebiet der Botanik, n.F. 1: 43-52.
Stosch, H.A. von (1982). On auxospore envelopes in diatoms. Bacillaria 5: 127-156.
Title Illustrations

| Scientific Name | Biddulphia |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | Frank E. Round |
| Life Cycle Stage | Vegetative phase |
| Body Part | theca |
| View | external, SEM |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann

|

| Scientific Name | Diatoms |
|---|---|
| Specimen Condition | Live Specimen |
| Life Cycle Stage | Vegetaive phase |
| View | girdle view, two cells |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann
|

| Scientific Name | Fallacia |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | David G. Mann |
| Life Cycle Stage | Vegetative phase |
| Body Part | frustule |
| View | exterior, SEM |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 David G. Mann
|
About This Page
This page is being developed as part of the Tree of Life Web Project Protist Diversity Workshop, co-sponsored by the Canadian Institute for Advanced Research (CIFAR) program in Integrated Microbial Biodiversity and the Tula Foundation.
David G. Mann
Royal Botanic Garden Edinburgh, United Kingdom
Correspondence regarding this page should be directed to David G. Mann at
Page copyright © 2008 David G. Mann
 Page: Tree of Life
Bacillariophytina.
Authored by
David G. Mann.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Bacillariophytina.
Authored by
David G. Mann.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 07 February 2010
Citing this page:
Mann, David G. 2010. Bacillariophytina. Version 07 February 2010 (under construction). http://tolweb.org/Bacillariophytina/125296/2010.02.07 in The Tree of Life Web Project, http://tolweb.org/

