



Caesalpinieae s.l. & s.s., Cassieae sensu stricto, Mimosoideae
Martin F. Wojciechowski



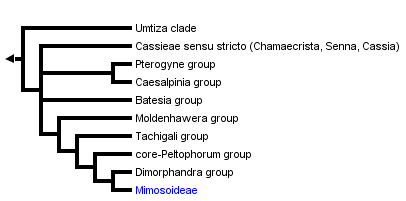


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The clade shown here, referred to as the "Caesalpinioid crown" clade (Wojciechowski et al., 2004), containing Cassieae subtribes Cassiinae and Ceratoniinae (sensu Irwin and Barneby, 1981) as well as tribe Caesalpinieae and the entire subfamily Mimosoideae, has been resolved in all molecular phylogenetic analyses (see below). The members of tribe Caesalpinieae have been divided into eight to nine informal generic groups (e.g., Polhill, 1994). These include the Umtiza, Pterogyne, Caesalpinia, Batesia, Moldenhawera, Tachigali, core-Peltophorum, and Dimorphandra groups, which as currently presented by Lewis (2005b), contain 56 genera and 430-440 species.
Discussion of Phylogenetic Relationships
Results from phylogenetic studies have led to the restructuring of some of the eight informal groups within Caesalpinieae. For example, recent analyses of molecular sequence data have supported some of these informal groups as monophyletic (e.g., Bruneau et al., 2001; Kajita et al., 2001; Wojciechowski et al. 2004) but the tribe as a whole is clearly paraphyletic, and the Mimosoideae and Cassieae sens. strict. are nested within its lineages. As a result of phylogenetic analysis, Bruneau et al. (2001) removed Ceratonia from Cassieae sensu lato (Lewis, 2005a), showing it was sister to Acrocarpus among the Caesalpinieae in the Umtiza clade, which now includes Gymnocladus, Gleditsia, Umtiza, Tetrapterocarpon, Arcoa, Acrocarpus, and Ceratonia (Herendeen et al. 2003).
The Caesalpinia group is a broad, apparently monophyletic group containing 20 or more genera (Lewis, 2005b) considered closely related to the type genus Caesalpinia. Numerous studies (e.g., Lewis and Schrire, 1995; Simpson and Lewis, 2003; Simpson et al., 2003) have clearly demonstrated that Caesalpinia, as traditionally circumscribed with c. 140 spp., is paraphyletic; Caesalpinia in the strict sense is now reduced to a genus of c. 25 spp. (Lewis, 2005b).
A recent molecular phylogenetic study by Marazzi et al. (2006) greatly clarified relationships within Senna. The Pterogyne and Moldenhawera groups contain only their name sakes, the genera Pterogyne and Moldenhawera, respectively. Haston et al. (2003) found that the Peltophorum group of Polhill (1994) was non-monophyletic and removed genera from the group limiting it to what is now referred to as the 'core-Peltophorum' group, with Schizolobium, Bussea, Peltophorum, Parkinsonia, Conzattia, Delonix, Covillea, and Lemuropisum. Relationships within this core-Peltophorum group have been further resolved by Haston et al. (2005).
With the exception of Dinizia (currently treated in Mimoseae), which appears more closely related to members of Caesalpinieae than to mimosoids on the basis of both molecular and morphological evidence (Luckow et al., 2003), the subfamily Mimosoideae is resolved as monophyletic with strong support (Luckow et al., 2003; Wojciechowski et al., 2004). Phylogenetic studies so far indicate that members of the Dimorphandra group of Caesalpinieae (e.g., Dimorphandra, Erythrophleum, Mora) comprise the sister group to the Mimosoideae, although relationships of lineages near the 'base' of the mimosoids clade are still poorly resolved.
References
Bruneau, A., F. Forest, P. S. Herendeen, B. B. Klitgaard, and G. P. Lewis. 2001. Phylogenetic relationships in the Caesalpinioideae (Leguminosae) as inferred from chloroplast trnL intron sequences. Systematic Botany 26: 487-514.
Haston, E. M., G. P. Lewis, and J. A. Hawkins. 2003. A phylogenetic investigation of the Peltophorum group (Caesalpinieae: Leguminosae). Pages 149-159 in Advances in Legume Systematics, part 10, higher level systematics (B. B. Klitgaard and A. Bruneau, eds.). Royal Botanic Gardens, Kew, UK.
Haston, E. M., G. P. Lewis, and J. A. Hawkins. 2005. A phylogenetic reappraisal of the Peltophorum group (Caesalpinieae: Leguminosae) based on the chloroplast trnL-F, rbcL, and rps16 sequence data. American J. Botany 92: 1359-1371.
Herendeen, P. S., A. Bruneau, G. P. Lewis. 2003. Phylogenetic relationships in caesalpinioid legumes: a preliminary analysis based on morphological and molecular data. Pages 37-62 in Advances in Legume Systematics, part 10, higher level systematics (B. B. Klitgaard and A. Bruneau, eds.). Royal Botanic Gardens, Kew, UK.
Herendeen, P. S., A. Bruneau, G. P. Lewis. 2003. Floral morphology in caesalpinioid legumes: testing the monophyly of the "Umtiza clade". International J. Plant Sciences 164 (5 Suppl.) S393-S407.
Irwin, H. S. and R. C. Barneby. 1981. Cassieae Bronn. Pages 97-106 in Advances in legume systematics, part 1 (R. M. Polhill and P. H. Raven, eds.). Royal Botanic Gardens, Kew, UK.
Kajita, T., H. Ohashi, Y. Tateishi, C. D. Bailey, and J. J. Doyle. 2001. rbcL and legume phylogeny, with particular reference to Phaseoleae, Millettieae, and Allies. Systematic Botany 26: 515-536.
Lewis, G. P. 2005a. Cassieae. Pages 111-125 in Legumes of the World (Lewis, G., B. Schrire, B. MacKinder, and M. Lock, eds.). Royal Botanic Gardens, Kew, UK.
Lewis, G. P. 2005b. Caesalpinieae. Pages 127-161 in Legumes of the World (Lewis, G., B. Schrire, B. MacKinder, and M. Lock, eds.). Royal Botanical Gardens, Kew, UK.
Lewis, G. P. and B. D. Schrire. 1995. A reappraisal of the Caesalpinia group (Caesalpinioideae: Caesalpinieae) using phylogenetic analysis. Pages 41-52 in Advances in Legume Systematics 7, phylogeny (M. D. Crisp and J. J. Doyle, eds.). Royal Botanic Gardens, Kew, UK.
Luckow, M., J. T. Miller, D. J. Murphy, and T. Livshultz. 2003. A phylogenetic analysis of the Mimosoideae (Leguminosae) based on chloroplast DNA sequence data. Pages 197-220 in Advances in Legumes Systematics, part 10, higher level systematics (B. B. Klitgaard and A. Bruneau, eds.). Royal Botanic Gardens, Kew, UK.
Marazzi, B., P. K. Endress, L. P. de Queiroz, and E. Conti. 2006. Phylogenetic relationships within Senna (Leguminosae, Cassinae) based on three chloroplast DNA regions: patterns in the evolution of floral symmetry and extrafloral nectaries. American J. Botany 93: 288-303.
Polhill, R. M. 1994. Classification of the Leguminosae. Pages xxxv - xlviii in Phytochemical dictionary of the Leguminosae (F. A. Bisby, J. Buckingham, and J. B. Harborne, eds.). Chapman and Hall, New York, NY.
Polhill, R. M., and J. E. Vidal. 1981. Caesalpinieae. Pages 81-95 in Advances in Legume Systematics, part 1 (R. M. Polhill and P. H. Raven, eds.). Royal Botanic Gardens, Kew, UK.
Simpson, B. B., L. L. Larkin, and A. Weeks. 2003. Progress towards resolving the relationships of the Caesalpinia group (Caesalpinieae: Caesalpinioideae: Leguminosae). Pages 123-148 in Advances in Legume Systematics, part 10, higher level sytematics (B. B. Klitgaard and A. Bruneau, eds.). Royal Botanic Gardens, Kew, UK.
Simpson, B. B., and G. P. Lewis. 2003. New combinations in Pomaria (Caesalpinioideae: Leguminosae). Kew Bulletin 58: 175-184.
Wojciechowski, M. F., M. Lavin, and M. J. Sanderson. 2004. A phylogeny of legumes (Leguminosae) based on analysis of the plastid matK gene resolves many well-supported subclades within the family. American J. Botany 91: 1846-1862.
Title Illustrations

| Scientific Name | Senna marilandica (L.) [=Cassia marilandica L.] |
|---|---|
| Location | Konza Prairie, Riley County, Kansas |
| Comments | Commonly called Wild Senna or Maryland Senna. |
| Specimen Condition | Live Specimen |
| Copyright | © Eva Horne |

| Scientific Name | Caesalpinia pulcherrima Sw. |
|---|---|
| Location | Arizona |
| Specimen Condition | Live Specimen |
| Copyright | © Chris Martin |

| Scientific Name | Ceratonia siliqua L. |
|---|---|
| Specimen Condition | Live Specimen |
| Copyright | © Annette Hoggemeier |
About This Page
Martin F. Wojciechowski

Arizona State University, Tempe, Arizona, USA
Correspondence regarding this page should be directed to Martin F. Wojciechowski at
Page copyright © 2006 Martin F. Wojciechowski
All Rights Reserved.
- First online 14 June 2006
- Content changed 14 June 2006
Citing this page:
Wojciechowski, Martin F. 2006. Caesalpinieae s.l. & s.s., Cassieae sensu stricto, Mimosoideae. Version 14 June 2006 (under construction). http://tolweb.org/Caesalpinieae_s.l._%26_s.s.%2C_Cassieae_sensu_stricto%2C_Mimosoideae/60239/2006.06.14 in The Tree of Life Web Project, http://tolweb.org/

