




Cupedidae
Thomas Hörnschemeyer




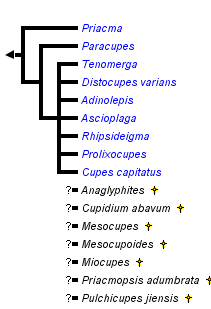


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
The Cupedidae represent the typical “reticulate” or “net-winged” beetles with incompletely sclerotized elytra that produce the characteristic reticulate appearance. Due to the long antennae and the general habitus these beetles otherwise resemble longhorn beetles (Cerambycidae).
The Cupedidae currently comprise 33 valid extant species. Since Neboiss’ (1984) revision of Cupes, nine genera are recognized. Three of these genera are monotypic with Priacma serrata (Leconte, 1861) and Cupes capitatus Fabricius, 1801 occurring in North America and Distocupes varians (Lea, 1902) in eastern Australia. Representatives of two more genera are present in the Australian region: The three species of Ascioplaga Neboiss, 1984 as well as the four species of Adinolepis Neboiss, 1984 are restricted to this region (Australia & New Caledonia). With 12 species, Tenomerga Neboiss, 1984 is the largest of the cupedid genera. Nine of them occur in the Asian and Oceanian regions, one (Tenomerga leucophaea (Newman, 1839)) in southern Africa and one (Tenomerga cinerea (Say, 1831)) in North America. The only other Archostemata to be found in the Ethiopian region belong to the genus Rhipsideigma Neboiss, 1984. One species (R. cretaceotincta (Kolbe, 1897)) is known from eastern Africa (Tanzania), the other four occur on Madagascar. The two remaining cupedid genera Paracupes Kolbe, 1898 and Prolixocupes Neboiss, 1960 occur in the Americas. In each genus there are two species. In Prolixocupes, one species lives in western South America and the other in western North America, whereas both species of Paracupes are found in South America.
The little available information on the biology and life cycle of Cupedidae is mainly based on North American and Australian species. Nevertheless, larvae have been described for Priacma serrata LeConte, 1861 (Ross & Pothecary 1970), Distocupes varians (Lea, 1902) (Neboiss 1968), Rhipsideigma raffrayi (Fairmaire, 1884) (Beutel & Hörnschemeyer 2002b), Tenomerga cinerea (Say, 1831) (Snyder 1913) and Tenomerga mucida (Chevrolat, 1829) (Fukuda 1938, 1939). However, beyond the description of usually one larval stage, only for T. cinerea and T. mucida a bit more information is available. Snyder (1913) was able to rear larvae of T. cinerea in the laboratory and he suggested that the species prefers wood infested with the fungus Daedalea quercina (Snyder 1956) as host substrate for its larvae. Fukuda (1941) reported a similar relationship with the fungus Stromatoscypha sp. (as Prorothelium sp.; Basidiomycetes: Schizophyllaceae) for T. mucida.
The fossil record of Cupedidae probably starts with fossils from the Jurassic (Ponomarenko 1966, 1969, 1997) that seem to be associated with the extant Priacma. Fossils that closely resemble Tetraphalerus and Omma are also known from Jurassic localities (Ponomarenko 1969). A comprehensive list of fossil Cupedidae can be found at the website "Classification of Fossil Beetles".
Taxonomic revisions of Cupedidae where published by Atkins (1963) and Neboiss (1960, 1984) who gave overviews of the species known at that time and their biogeography. Nebois (1960, 1984) did a comprehensive rearrangement of the species then summarized under the genus Cupes. He described six new genera and left Cupes capitatus as the only species in the genus.
Characteristics
With the usually slender body and with antennae nearly as long as the body, Cupedidae resemble longhorn beetles (Cerambycidae). Nevertheless, this resemblance is really only supeficial and disappears when one looks at a cupedid from the side, since most Cupedidae (Priacma serrata might be an exception) are conspicuously dorsoventrally flattened.
Very characteristic features of Cupedidae are the more or less pronounced dorsal head protuberances. They are present in all species but are smallest in Priacma, bulbous in Prolixocupes and tall and steep in Rhipsideigma and Cupes.
The colors are usually in the range brown, gray, black, with a few exceptions like the orange head of Cupes capitatus and the black head and prothorax with bright orange markings in Rhipsideigma lugubris. In all Cupedidae the visible colors and color patterns are caused by scales that densely cover the uniformly dark brown to black cuticle of these beetles.
Discussion of Phylogenetic Relationships
Analyses of the phylogenetic relationships of Cupedidae, based on morphological characters, were published by Hörnschemeyer et al. (2002, 2006), Beutel & Hörnschemeyer (2002a, 2002b), Beutel et al. (2008), Friedrich et al. (2008) with most of the genera represented by one species. Hörnschemeyer (2009) published a species level phylogeny of Archostemata also based on morphology and comprising 37 of 45 known extant species. The relationships of cupedid genera reconstructed in all these publications are quite similar. The differences that occur might be attributed mainly to the often sparse taxon sampling.
In general Cupedidae seem to be composed of a few monophyletic groups with a clade comprising Priacma and Paracupes as the sistergroup of all other Cupedidae. The next branch is formed by Prolixocupes and then follow the “Tenomerga-like” species. Within this group the phylogeny seems to mirror the biogeography of the species: an Australian/New Caledonian clade contains Adinolepis and Ascioplaga, one clade with African, European (from Baltic amber and Pliocene sediments) and American species comprises Cupes, Rhipsideigma and Tenomerga leucophaea (Newman, 1839) and the last group is composed of the remaining eastern Asian and North American species of Tenomerga. In-between the Australian and the Eurasian groups is the lonely Distocupes varians (Lea, 1902), which might be the sistergroup of the Australian clade (Hörnschemeyer et al. 2006) or of the Eurasian group (Hörnschemeyer 2009).
Click on an image to view larger version & data in a new window 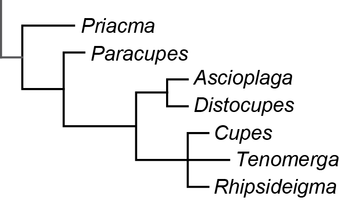 Relationships of some genera of Cupedidae as reconstructed by Beutel et al. (2008) | Click on an image to view larger version & data in a new window 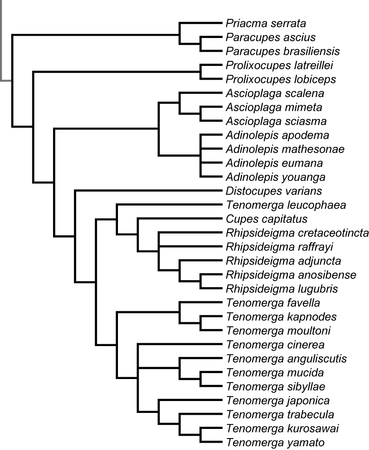 Relationships of species of Cupedidae as reconstructed by Hörnschemeyer (2009) |
An interesting but probably not completely unexpected result of the species level analysis of Hörnschemeyer (2009) was that Tenomerga probably is not a monophyletic group. Tenomerga leucophaea turned out to be a part of a monophylum comprising Cupes capitatus and the genus Rhipsideigma but no other Tenomerga species. Already Nebois (1984: 457) recognized that T. leucophaea is different from the other species of Tenomerga in a number of characters but nevertheless put it in this newly established genus. The phylogenetic analysis revealed that, based on characters of the head, the mouthparts and the mail genitalia, T. leucophaea is more closely related to Rhipsideigma and Cupes than to the remaining species of Tenomerga. If this result should be confirmed e.g. by an analysis based on genetic data, a taxonomic revision of the concerned genera would be justified and necessary.
Another interesting suggestion from the analysis of Hörnschemeyer (2009) is a possible close relationship of the enigmatic Crowsoniella relicta Pace, 1975 with Cupedidae. Nevertheless, this interpretation is nearly exclusively based on characters of the male genitalia and needs to be checked with other character systems.
References
Atkins, M. D. 1963. The Cupedidae of the world. Canadian Entomologist 95: 140-162.
Beutel, R. G., Ge, S.-Q. & Hörnschemeyer, T. 2008. On the head morphology of Tetraphalerus, the phylogeny of Archostemata and the basal branching events in Coleoptera. Cladistics 24: 270-298.
Beutel, R. G. & Hörnschemeyer, T. 2002a. Larval morphology and systematic position of Micromalthus debilis LeConte, 1878 (Coleoptera, Micromalthidae). Systematic Entomology 27: 169-190.
Beutel, R. G. & Hörnschemeyer, T. 2002b. Description of the larva of Rhipsideigma raffrayi (Coleoptera, Archostemata), with phylogenetic and functional implications. European Journal of Entomology 99: 53-66.
Carpenter, F. M. 1992. Treatise on Invertebrate Paleontology. Part R. Arthropoda 4, Volume 3: Superclass Hexapoda. Geological Society of America and University of Kansas, Boulder, Colorado and Lawrence, Kansas.
Crowson, R.A. 1962. Observations on the beetle family Cupedidae, with descriptions of two new fossil forms and a key to the recent genera.. Annals & Magazine of Natural History 5, ser. 13, No. 51: 147-157.
Friedrich, F., Farell, B.D., Beutel, R.G., 2008. The thoracic morphology of Archostemata and the relationships of the extant suborders of Coleoptera (Hexapoda). Cladistics 25: 1-37.
Fukuda, A 1938. Description of the larva and pupa of Cupes clathratus. Transactions of the natural History Society of Formosa, Taihoku. 28: 390-393.
Fukuda, A. 1939. On the larva of Cupes clathratus. Transactions of the natural History Society of Formosa, Taihoku. 29: 75-82.
Fukuda, A. 1941. Biology of Tenomerga mucida. Transactions of the Natural History Society of Formosa 31: 394-399.
Hörnschemeyer, T., 2009. The species-level phylogeny of archostematan beetles - where do Micromalthus debilis and Crowsoniella relicta belong? Systematic Entomology 34(3): 533-558.
Hörnschemeyer, T., Beutel, R. G. & Pasop, F. 2002. Head Structures of Priacma serrata Leconte (Coleptera, Archostemata) Inferred from X-ray Tomography. Journal of Morphology 252(3): 298-314.
Hörnschemeyer, T., Goebbels, J., Weidemann, G., Faber, C., Haase, A., 2006. The head of Ascioplaga mimeta (Coleoptera: Archostemata) and the phylogeny of Archostemata. European Journal of Entomology 103: 409-423.
Laporte, F.. L. de 1836. Etudes entomologiques, ou descriptions d'insectes nouveaux et observations sur la synonymie. Revue entomologique, publie´e par G. Silbermann. IV: 5-60.
Lawrence, J. F. & Newton, A. F. 1995. Families and subfamilies of Coleoptera. In: Pakaluk & Slipinski: Biology, Phylogeny, and Classification of Coleoptera. Vol. 2. Muzeum i Instytut Zoologii PAN, Warzawa: 779-1006.
Lea, A. M. 1902. Description of new species of Australian Coleoptera, Part VI. The Proceedings of the Linnean Society of New South Wales 26: 485-487 & 513.
Neboiss, A. 1960. On the family Cupedidae, Coleoptera. Proceedings of the Royal Society of Victoria 72(1): 12-20.
Neboiss, A. 1968. Larva and pupa of Cupes varians Lea, and some observations on ist biology (Coleoptera: Cupedidae). Memoirs of the National Museum of Victoria 28: 17-19.
Neboiss, A. 1984. Reclassification of Cupes Fabricius (s.lat.), with descriptions of new genera and species (Cupedidae: Coleoptera). Systematic Entomology 9: 443-477.
Newman, E. 1839. XXXV.-Supplementary note to the synonymy of Passandra. Annals and Magazine of Natural History 3 (18): 303-305.
Pace, R. (1975). An exceptional endogeous beetle: Crowsoniella relicta n. gen. n. sp. of Archostemata Tetraphaleridae from Central Italy. Bolletino del Museo civico di Storia naturale Verona 2: 445-458.
Ponomarenko, A. G. 1966. New beetles of the Family Cupedidae (Coleoptera) from Mesozoic deposits of Transbaikalia. Revue d'Entomologie de l'URSS 45:138-143.
Ponomarenko, A. G. 1969. Historical development of the Coleoptera-Archostemata. Trudy paleontologicheskogo instituta akademii nauk SSSR 125:1-240.
Ponomarenko, A. G. 1972. Erster fund eines Cupediden im neogen Europas (Coleoptera: Cupedidae). Vestnik Cs. spol. zool. 37(2):101-103.
Ponomarenko, A. G. 1997. New beetles of the Family Cupedidae from the Mesozoic of Mongolia. Ommatini, Mesocupedini, Priacmini. Paleontological Journal 31(4):389-399.
Ross, D. A. & Pothecary, D. D. 1970. Notes on adults, eggs, and firts-instar larvae of Priacma serrata (Coleoptera: Cupedidae). The Canadian Entomologist 102: 346-348.
Snyder, T. E. 1913. Record of the rearing of Cupes concolor Westw. (Coleoptera). Proceedings of the Entomologica Society of Washington 15: 30-31.
Snyder, T. E. 1956. Cupes and Daedalea. The Coleopterists Bulletin: An International Journal Devoted to the Study of Beetles 10 (2): 20.
Information on the Internet
- Classification of Fossil Beetles. A. G. Ponomarenko and A. G. Kirejtshuk, Zoological Institute of the Russian Academy of Sciences.
Title Illustrations

| Scientific Name | Priacma serrata |
|---|---|
| Location | Oregon, U.S.A. |
| Image Use |
 This media file is licensed under the Creative Commons Attribution License - Version 3.0. This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1995 David R. Maddison

|

| Scientific Name | Tenomerga cinerea |
|---|---|
| Specimen Condition | Dead Specimen |
| Life Cycle Stage | Adult |
| View | Dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1996 David R. Maddison
|

| Scientific Name | Rhipsideigma raffrayi |
|---|---|
| Location | Madagascar |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1995 David R. Maddison
|

| Scientific Name | Prolixocupes latreillei |
|---|---|
| Specimen Condition | Dead Specimen |
| Life Cycle Stage | Adult |
| View | Dorsal |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1996 David R. Maddison
|
About This Page
University Göttingen, Institute for Zoology & Anthropology, Dept. Morphology & Systematics
Correspondence regarding this page should be directed to Thomas Hörnschemeyer at
Page copyright © 2011
 Page: Tree of Life
Cupedidae .
Authored by
Thomas Hörnschemeyer.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Cupedidae .
Authored by
Thomas Hörnschemeyer.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- Content changed 23 December 2010
Citing this page:
Hörnschemeyer, Thomas. 2010. Cupedidae . Version 23 December 2010. http://tolweb.org/Cupedidae/8999/2010.12.23 in The Tree of Life Web Project, http://tolweb.org/

