



Mysmenidae
Lara Lopardo and Gustavo Hormiga


The family is currently undergoing taxonomic recircumscription by Lopardo and Hormiga (in prep.). It includes 107 described species in 21 genera.
- Anjouanella
- Brasilionata
- Calodipoena
- Calomyspoena
- Crassignatha
- Iardinis
- Isela
- Itapua
- Kekenboschiella
- Kilifina
- Leviola
- Maymena
- Microdipoena
- Mysmena
- Mysmenella
- Mysmeniola
- Mysmenopsis
- Phricotelus
- Tamasesia
- Taphiassa
- Trogloneta
Introduction
Mysmenids are one of the least studied groups of orb-weaving spiders, mainly because of their minuscule size (0.6-3 mm; Fig. 1) and cryptic life history. The family Mysmenidae is distributed worldwide, and with only around 100 described species, the diversity of mysmenids is clearly under-sampled as numerous undescribed species of this family have been collected and/or exist in museum collections around the world.
Mysmenids live mainly in leaf litter and other cryptic places in humid habitats. Mysmenid web-spinning species seem to usually prefer the interstices of leaf litter and small cavities about 5-15 cm in diameter (depending on the size of the spider) created by the top layer of leaves. They can be collected by beating foliage, using pitfall traps, Berlese funnels, Winkler devices (e.g. Wheeler and McHugh, 1987), or just manually.
Only a few mysmenid spiders have been documented from the fossil record (eight species in five genera). Seven fossil mysmenids have been described from Tertiary ambers from the Miocene (15–20 Ma; two species from Dominican amber), Miocene–Oligocene (19–27 Ma, one species from Chiapas amber), and Eocene (44 Ma, two species from the Baltic amber, and two species from the Baltic and Bitterfeld ambers). A relatively recent species was described from Madagascan copal (a semi-fossilized resin less than two million years old).



Figure 1. Representatives of Mysmenidae.
Left: Mysmena tasmaniae female. Center: Microdipoena nyungwe male. Right: Maymena ambita male. Composite digital images © Lara Lopardo.
Characteristics
Mysmenids resemble some theridiids in their somatic morphology, as well as some members of the more closely related symphytognathoid families (Anapidae, Symphytognathidae, Theridiosomatidae and Synaphridae).Male mysmenids can be distinguished from other araneoids by the presence of a metatarsal clasping spine on the first legs (Fig. 2) and by a particular composition and arrangement of their copulatory organs (palps, Fig. 3). Females have a distinct dark modification, like a spot, on the apical ventral surface of at least the first femur (Fig. 4, sometimes on the second femur and in some species such modification is also present in males). Both sexes of mysmenids have the anterior median eyes on a protruding area (or all eyes on tubercle); a prolateral row of modified setae on the first tarsus (Fig. 2); a distinctly thicker and curved seta on the chelicerae; and denticles in the cheliceral fang furrow (Fig. 5).


Figure 2. Features of the first leg of a male of Microdipoena nyungwe. Left: Prolateral clasping spines. Right: Prolateral tarsal row of modified setae. SEM images © Lara Lopardo
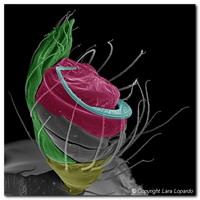
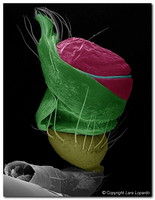
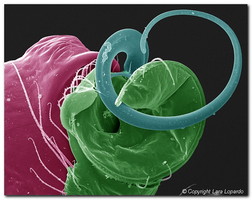
Figure 3. Left palps of male mysmenids (Cymbium depicted in green, Embolus in blue and Conductor in red). Left, center: Two unidentified Mysmena species; Right: Microdipoena nyungwe. SEM images © Lara Lopardo


Figure 4. Modifications of the first femur in female Calodipoena incredula (left) and Mysmenopsis dipluramigo (right). SEM images © Lara Lopardo

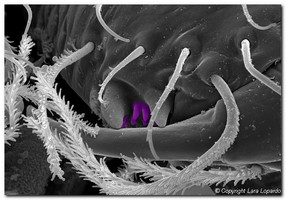
Figure 5. Denticles (highlighted in purple) in the cheliceral fang furrow of Maymena rica (left) and an unidentified mysmenid (right). SEM images © Lara Lopardo
Very little is known about the biology and natural history of mysmenids. Eleven species in three mysmenid genera have been reported to be kleptoparasites on the webs of other spiders, such as diplurids, tengellids, or lycosoids. The webs of most other mysmenids have never been documented. Mysmenids are known to build two main types of web architecture. Mysmena and Microdipoena species build highly modified three-dimensional spherical-shaped orb webs. In contrast, the webs of Maymena species are mainly planar but the central hub is distorted upwards by one to several radial lines that attach to substrate above the web. Members of the symphytognathoid family Anapidae build webs identical to the ones built by Maymena species.





Figure 6. Mysmenid webs. Top left: Mysmena tasmaniae (Queensland, Australia). Top center: female Mysmena sp. with eggsac (Misiones, Argentina). Top right: Mysmena sp. (Chiapas, Mexico), peripheral radial lines and sticky silk removed to expose hub. Bottom left: Maymena sp. (Misiones, Argentina). Bottom right: Same, detail to hub. Images © Lara Lopardo
Discussion of Phylogenetic Relationships
After more than 80 years since the erection of the family, the first generic-level cladistic analysis of Mysmenidae is in progress (Lopardo and Hormiga, in prep.). This combined analysis using morphological and molecular data will test the monophyly of the family and its genera, and will also hypothesize a placement of Mysmenidae within the Symphytognathoidea. Until now the placement of Mysmenidae within this group has been based exclusively on morphology and behavior. Two cladistic analyses included mysmenids in the past (Griswold et al., 1998; Schütt 2003) and, to a limited extent, tested the monophyly of the family. These two analyses included only two mysmenid representatives. Both placed the Mysmenidae within Symphytognathoidea, either as sister to Symphytognathidae, or to a clade comprising Anapidae plus Symphytognathidae. More recently, a new phylogenetic hypothesis for Araneoidea and Symphytognathoidea has been proposed (Lopardo and Hormiga, 2008), revising and expanding some of the previous phylogenetic work. Lopardo and Hormiga (2008) placed Mysmenidae either as sister to the clade comprising Anapidae and Symphytognathidae (as originally proposed by Griswold et al., 1998) or as sister to Theridiosomatidae. Only two molecular phylogenetic analyses have included mysmenid representatives, as part of the outgroup taxa (Arnedo et al., 2004; Rix et al., 2008).
References
Arnedo, M, Coddington, J, Agnarsson, I and Gillespie, R. 2004. From a comb to a tree: Phylogenetic relationships of the comb-footed spiders (Araneae, Theridiidae) inferred from nuclear and mitochondrial genes. Molecular Phylogenetics and Evolution 31: 225-245.
Baert, L. 1984a. Spiders (Araneae) from Papua New Guinea. IV. Ochyroceratidae, Telemidae, Hadrotarsidae and Mysmenidae. Indo-Malayan Zoology 2: 225-244.
Baert, L. 1984b. Mysmenidae and Hadrotarsidae from the Neotropical Guaraní zoogeographical province (Paraguay and south Brasil) (Araneae). Revue Suisse De Zoologie 91: 603-616.
Baert, L. 1986. Mysmenidae from the Comoro Islands (Araneae). Revue de Zoologie Africaine 100: 265-267.
Baert, L. 1988. The Ochyroceratidae and Mysmenidae from Sulawesi (Araneae). Indo-Malayan Zoology 5: 9-22.
Baert, L. 1990. Mysmenidae (Araneae) from Peru. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique Entomologie 60: 5-18.
Baert, L and Maelfait, J-P. 1983. Spiders of the Galápagos Islands. I. Mysmenidae (Araneae). Bulletin of the British Arachnological Society 6: 102-108.
Brescovit, A and Lopardo, L. 2008. The first record on the spider genus Trogloneta Simon in the southern hemisphere (Araneae, Mysmenidae), with descriptions of three new species from Brazil and remarks on the morphology. Acta Zoologica (Stockholm) 89: 93106.
Brignoli, P. 1970. Contribution à la connaissance des Symphytognathidae paléarctiques (Arachnida, Araneae). Bulletin du Museum National d'Histoire Naturelle, Paris 41: 1403-1420.
Brignoli, P. 1980. On few Mysmenidae from the Oriental and Australian regions (Araneae). Revue Suisse De Zoologie 87: 727-738.
Coddington, J. 1986b. The monophyletic origin of the orb web. In: Spider Webs, Behavior and Evolution. (WA Shear, e). Stanford University Press.319-363
Eberhard, W. 1987. Orb webs and construction behavior in Anapidae, Symphytognathidae, and Mysmenidae. Journal of Arachnology 14(3): 339-356.
Forster, R. 1959. The spiders of the family Symphytognathidae. Transactions of the Royal Society of New Zealand 86: 269-329.
Forster, R and Platnick, N. 1977. A review of the spider family Symphytognathidae (Arachnida, Araneae). American Museum Novitates 2619: 1-29.
Gertsch, W. 1960. Descriptions of American spiders of the family Symphytognathidae. American Museum Novitates 1981: 1-40.
Griswold, C. 1985. Isela okuncana, a new genus and species of kleptoparasitic spider from southern Africa (Araneae: Mysmenidae). Annals of the Natal Museum 27: 207-217.
Griswold, C, Coddington, J, Hormiga, G and Scharff, N. 1998. Phylogeny of the orb-web building spiders (Araneae, Orbiculariae: Deinopoidea, Araneoidea). Zoological Journal of the Linnean Society 123: 1-99.
Hajer, J. 2000. Web of Trogloneta granulum Simon (Araneae, Mysmenidae). Bulletin of the British Arachnological Society 11: 333-338.
Hajer, J. 2002. The spinning activity of the spider Trogloneta granulum Simon, 1922 (Araneae, Mysmenidae). Acta Universitatis Purkynianae Studia Biologica VI: 21-38.
Hajer, J and Reháková, D. 2003. Spinning activity of the spider Trogloneta granulum (Araneae, Mysmenidae): web, cocoon, cocoon handling behaviour, draglines and attachment discs. Zoology 106(3): 223-231.
Hickman, V. 1979. Some Tasmanian spiders of the families Oonopidae, Anapidae and Mysmenidae. Papers and Proceedings of the Royal Society of Tasmania 113: 53-79.
Kraus, O. 1967. Zur Spinnenfauna Deutschlands, II. Mysmena jobi n. sp, eine Symphytognathide in Mitteleuropa (Arachnida: Araneae: Symphytognathidae). Senckenbergiana Biologica 48: 387-399.
Levi, H. 1956. The spider genus Mysmena in the Americas (Araneae, Theridiidae). American Museum Novitates 1801: 1-13.
Lopardo, L and Coddington, J. 2005. Chapter 40: Mysmenidae. In: Spiders of North America: an identification manual. (Ubick, D, Cushing, P and Paquin, P). American Arachnology Society.175-177
Lopardo, L, Dupérré, N and Paquin, P. 2008. Expanding horizons... The first report of the genus Mysmena (Araneae, Mysmenidae) from continental North America, with the description of a new species. Zootaxa 1718: 3644.
Penney, D. 2000. Miocene spiders in Dominican amber (Oonopidae, Mysmenidae). Palaeontology 43: 343-357.
Petrunkevitch, A. 1971. Chiapas amber spiders, II. University of California Publications in Entomology 63: 1-44.
Platnick, N. 2009. The world spider catalog, version 9.5. American Museum of Natural History, online at http://research.amnh.org/entomology/spiders/catalog/index.html.
Platnick, N and Shadab, M. 1978. A review of the spider genus Mysmenopsis (Araneae, Mysmenidae). American Museum Novitates 2661: 1-22.
Rix, M, Harvey, M and Roberts, J. 2008. Molecular phylogenetics of the spider family Micropholcommatidae (Arachnida: Araneae) using nuclear rRNA genes (18S and 28S). Molecular Phylogenetics and Evolution 46(3): 1031-1048.
Schütt, K. 2003. Phylogeny of Symphytognathidae s.l. (Araneae, Araneoidea). Zoologica Scripta 32: 129-151.
Thaler, K. 1975. Trogloneta granulum Simon, eine weitere Reliktart der Nordostalpen (Arachnida, Aranei, "Symphytognathidae"). Revue Suisse De Zoologie 82: 283-291.
Thaler, K. 1995. Mysmeniola spinifera n.gen. n.sp., eine merkwürdige Kleinspinne aus Venezuela (Arachnida, Araneae: Mysmenidae). Mitteilungen der Schweizerischen Entomologischen Gesellschaft 68: 429-433.
Wheeler, Q.D., and J.V. McHugh. 1987. A portable and convertible Moczarski/Tullgren extractor for fungus and litter coleoptera. Coleopterist Bulletin 4: 912.
Wunderlich, J. 1998. Beschreibung der ersten fossilen Spinnen der Unterfamilien Mysmeninae (Anapidae) und Erigoninae (Linyphiidae) im Dominikanischen Bernstein (Arachnida: Araneae). Entomologische Zeitschrift, Frankfurt am Main 108: 363-367.
Wunderlich, J. 2004. The fossil spiders of the family Anapidae s. l. (Aeaneae) [sic] in Baltic, Dominican and Mexican amber and their extant relatives, with the description of the new subfamily Comarominae. Beiträge zur Araneologie 3: 1020-1111.
Title Illustrations

| Scientific Name | Anjouanella comorensis |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | Lara Lopardo |
| Sex | Female |
| Life Cycle Stage | adult |
| Body Part | habitus |
| View | lateral |
| Collection | MRAC |
| Type | paratype |
| Copyright |
© 2009

|

| Scientific Name | Anjouanella comorensis |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | Lara Lopardo |
| Sex | Male |
| Life Cycle Stage | adult |
| Body Part | habitus |
| View | lateral |
| Collection | MRAC |
| Type | holotype |
| Copyright |
© 2009
|
About This Page
George Washington University, Washington, D. C., USA
Gustavo Hormiga
George Washington University, Washington, D. C., USA
Correspondence regarding this page should be directed to Lara Lopardo at and Gustavo Hormiga at
Page copyright © 2009 and Gustavo Hormiga
 Page: Tree of Life
Mysmenidae.
Authored by
Lara Lopardo and Gustavo Hormiga.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Mysmenidae.
Authored by
Lara Lopardo and Gustavo Hormiga.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 07 December 2006
- Content changed 10 March 2009
Citing this page:
Lopardo, Lara and Gustavo Hormiga. 2009. Mysmenidae. Version 10 March 2009 (under construction). http://tolweb.org/Mysmenidae/2803/2009.03.10 in The Tree of Life Web Project, http://tolweb.org/


