



Osteostraci
Philippe Janvier

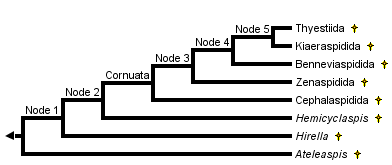


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxThe main characteristics supporting the nodes of this phylogeny are:
- Node 1: Narrow-based paired fins, oralobranchial cavity closed ventrally by large dermal platelets
- Node 2: Anterior dorsal fin reduced to a large median dorsal scute
- Cornuata: Cornual processes
- Node 3: Large median dorsal scute lost.
- Node 4: First canal leading from the inner ear to lateral fields branches close to the orbit.
- Node 5: Posterior ventral myodome of the orbit (for recti eye muscles) reduced.
Introduction
The Osteostraci, or osteostracans, are a major clade (about 200 species) of fossil, armored jawless vertebrates which lived from the Early Silurian (about 430 million years) to the Late Devonian (about 370 million years). Most of them have a characteristic horseshoe-shaped head, which consists of a massive endoskeletal skull, covered with a shield of dermal bone. On the dorsal surface of the head are the closely-set eyes, a pineal foramen, and a median, keyhole-shaped nasohypophysial opening. In addition, there are peculiar "fields" (in fact, depressions of the braincase, covered with loose platelets of dermal bone), which have been regarded as either sense organs or electric organs. The mouth and gill openings are, like in the Galeaspida, situated on the ventral side of the head. Osteostracans also have large, pad-shaped paired fins. Most osteostracans are about 20 to 40 cm. in total length, but some species could be extremely small (about 4 cm in length). The largest species is about one metre in length.

The zenaspidid osteostracan Zenaspis pagei, from the Lower Devonian of Scotland.
Osteostracans, also known as "cephalaspids", played an important role in the history of vertebrate palaeontology, as they were the first fossil jawless fishes whose internal anatomy has been described in detail, thereby raising heated debates about the organization of the primitive vertebrate head.
Most osteostracans lived in marginal, marine environments, some of them being regarded as fresh water. Their horseshoe-shaped headshield suggests a benthic mode of life, but it is possible that some of them, such as the Boreaspididae or the Kiaeraspidida were more active swimmers. They are most diverse in the Early Devonian and become very rare in the Middle and Late Devonian. Osteostracans are widely distributed in North America, Europe, Siberia, and Central Asia, north of the Tian-Shan.
Characteristics
Osteostracans are characterized by:
- Two lateral and one median depression in the dorsal surface of the endoskeletal headshield, referred to as "cephalic fields". These fields are covered by polygonal, exoskeletal platelets and are connected to the inner ear by means of large branching canals. They have been variously interpreted as having housed electric organs, expansions of the labyrinth, or special organs of the sensory-line system. To date, their function remains unknown.
- A peculiar, leaf-shaped, horizontal fin underlying the caudal fin. This may be a modified anal fin.
Osteostracans resemble the Galeaspida and Pituriaspida in having a massive endoskeletal headshield, made up by cartilage (sometimes with globular calcifications) lined with a thin layer of cellular perichondral bone. As in the latter two groups, this endoskeletal headshield is hollowed ventrally by a large oralobranchial cavity, the roof of which shows impressions of the gills and velum. It is undecided whether this headshield only represents the braincase, or also includes part of the visceral and scapular skeleton. This endoskeletal headshield is covered with an exoskeletal headshield, generally made up by polygonal platelets of cellular bone and bearing tubercles of mesodentine. The oralobranchial cavity is closed ventrally by either minute scales or larger dermal platelets. The mouth and gill openings (8 to 10) open ventrally, along the rim of the oralobranchial cavity. Dorsally, the headshield is pierced by two closely-set orbits, between which opens a pineal foramen (often piercing a small, separate plate), and a median, keyhole-shaped nasohypophysial opening. The nasohypophysial opening is the external opening of the small olfactory organ and the blind hypophysial tube. This remarkable similarity to lampreys has long suggested that the two groups were close relatives and were included, along with the Anaspida in a clade Cephalaspidomorphi. Current theories of craniate interrelationships suggest, however, that this characteristic is homoplastic.
Thanks to the perichondral ossification of the endoskeletal headshield, the internal anatomy of osteostracans could be studied in great details. It shows the brain cavity, with a paired recess for the cerebellum, the orbits, with myodomes for the eye muscles, the labyrinth cavity, with two vertical semicircular canals, and the canals for the cranial nerves and blood vessels. There have been heated debates about the path of the cranial nerves, in particular those innervating the gill arches. Some considered that the maxillary branch of the trigeminus nerve innervated a premandibular gill arch. Others considered that the gill anatomy of osteostracans was similar to that of larval lampreys, the foremost gill being that of the hyoid arch. The most intriguing feature of osteostracan anatomy is the large, branching canals which lead from the labyrinth to the lateral and dorsal cephalic "fields". Some interpret then as expansions of the membranous labyrinth, containing endolymph, whereas others regard them as having housed either sensory or electromotor nerves.
Osteostracans have paired fins which contained a musculature and were attached to the posterolateral region of the endoskeletal headshield.
The body of osteostracans is massive, covered with large, diamond-shaped scales, and bears primitively two dorsal fins (reduced to one or none in the more advanced forms). The tail is epicercal (i.e. the lobe containing the notochord points posterodorsally, as in sharks). The caudal fin web is underlain by a peculiar horizontal, leaf-shaped, fin web, which may represent a modified anal fin. Sensory-line canals are developed on the head and part of the body.Discussion of Phylogenetic Relationships
The Osteostraci include a major clade, the Cornuata, characterized by pointed lateral processes of the headshield, in front of the paired fins (homoplastic with the cornual processes of the Galeaspida and Pituriaspida), and a few primitive genera (Ateleaspis, Aceraspis, Hirella, Hemicyclaspis) which represent "basal" osteostracans. The Early Silurian Ateleaspis is regarded as the most generalized osteostracan.The Cornuata comprise five major taxa: the Cephalaspidida, Zenaspidida, Benneviaspidida, Kiaeraspidida, and Tyestiida. There are a few other minor taxa of uncertain affinities, such as the Tannuaspidida from Central Asia (Tuva).
The Cephalaspidida are characterized by broad and flattened cornual processes, onto which extend the lateral fields. Cephalaspis is the classical representative of this group which, however, includes many other forms, such as the large Parameteoraspis, whose headshield can reach 45 cm in breadth.
The Zenaspidida have a more massive headshield, often bearing a posterior median crest or spinal process. They have a characteristic ornamentation of large tubercles, surrounded by smaller ones.
The Benneviaspidida have a flattened headshield and their exoskeleton has lost the polygonal pattern of the platelets, to form a continuous layer. Some benneviaspidids, such as the Boreaspididae, develop a long rostral process, like some galeaspids and pituriaspids. One of them, Tauraspis, is unique in having a pair of forward pointing "horns".
The Kiearaspidida are small osteostracans whose headshield extends posteriorly into a long abdominal division and whose cornual processes are extremely reduced or lost. Some of them, the Acrotomaspididae, are very specialized forms, with extremely reduced cephalic fields and their mouth opens anterodorsally.
The Thyestiida are also small osteostracans, characterized by the special structure of their sensory-line canals. They include forms, the Tremataspididae, which have lost the paired fins and cornual process, their headshield becoming olive-shaped. This may represent an adaptation to burrowing habits.
Among all "ostracoderms", osteostracans (possibly along with pituriaspids) share the largest number of uniquely derived characteristics with the gnathostomes: cellular bone, sclerotic ring in eyes, paired fins containing musculature, and epicercal caudal fin.
References
Adrain, J. M. and Wilson, M. V. H. (1994). Early Devonian cephalaspids (Vertebrata: Osteostraci: Cornuata) from the southern Mackenzie Mountains, N.W.T., Canada. Journal of Vertebrate Paleontology, 14, 301-319.
Afanassieva, O. (1991). (The cephalaspids of Soviet Union). Trudy Paleontologicheskogo Instituta Akademia Nauk, 248, Moscow. (In Russian.)
Afanessieva, O. and Janvier, P. (1985). Tannuaspis, Tuvaspis and Ilemoraspis, endemic osteostracan genera from the Silurian and Devonian of Tuva and Khakassia (USSR). Géobios, 18, 493-505.
Janvier, P. (1981). Norselaspis glacialis n.g., n.sp. et les relations phylogénétiques entre les Kiaeraspidiens (Osteostraci) du Dévonien inférieur du Spitsberg. Palaeovertebrata, 11(2-3), 19-131.
Janvier, P. (1984). The relationships of the Osteostraci and Galeaspida. Journal of Vertebrate Paleontology, 4, 344-358.
Janvier, P. (1985a). Les Céphalaspides du Spitsberg. Anatomie, phylogénie et systématique des Ostéostracés siluro-dévoniens. Révision des Ostéostracés de la Formation de Wood Bay (Dévonien inférieur du Spitsberg). Cahiers de Paléontologie, Centre national de la Recherche scientifique, Paris.
Janvier, P. (1985b). Les Thyestidiens (Osteostraci) du Silurien de Saaremaa (Estonie). Première partie: Morphologie et anatomie, Annales de Paléontologie, 71(2), p.83-147. Deuxième partie: Analyse phylogénétique, répartition stratigraphique, remarques sur les genres Auchenaspis, Timanaspis, Tyriaspis, Didymaspis, Sclerodus et Tannuaspis. Annales de Paléontologie, 71(3), p.187-216.
Janvier, P. (1996). Early vertebrates. Oxford Monographs in Geology and Geophysics, 33, Oxford University Press, Oxford.
Mark-Kurik, E. & Janvier, P. (1995). Early Devonian osteostracans from Severnaya Zemlya, Russia. Journal of Vertebrate Paleontology. 15(3):449-462.
Ritchie, A. (1967). Ateleaspis tessellata Traquair, a non-cornuate cephalaspid from the Upper Silurian of Scotland. Zoological Journal of the Linnean Society, 47, 69-81.
Stensiö, E. A. (1927). The Devonian and Downtonian vertebrates of Spitsbergen. 1. Family Cephalaspidae. Skrifer om Svalbard og Ishavet, 12, 1-391.
Stensiö, E.A. (1964). Les Cyclostomes fossiles ou Ostracodermes. In: Traité de paléontologie (ed. J. Piveteau), Vol. 4(1), pp. 96-383, Masson, Paris
Wängsjö, G. (1952). The Downtonian and Devonian vertebrates of Spitsbergen. 9. Morphologic and systematic studies of the Spitsbergen cephalaspids. Results of Th. Vogt's Expedition 1928 and the English-Norwegian-Swedish Expedition in 1939. Norsk Polarinstitutt Skrifter, 97, 1-611.
Watson, D. M. S. (1954). A consideration of ostracoderms. Proceedings of the Royal Society of London, B, 238, 1-25.
White, E. I. and Toombs, H. A. (1983). The cephalaspids from the Dittonian at Cwm Mill, near Abergavenny, Gwent. Bulletin of the British Museum (natural History), Geology, 37, 149-171.
Whiting, H. P. (1977). Cranial nerves in lampreys and cephalaspids. In Problems in Vertebrate Evolution, Linnean Society Symposium Series, No. 4, (ed. S.M. Andrews, R. S. Miles, and A.D. Walker), pp. 1-23. Academic Press, London.
Title Illustrations

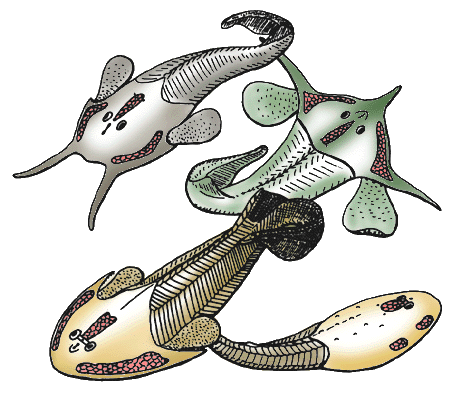
| Scientific Name | Tauraspis, Hoelaspis, Tremataspis, Zenaspis |
|---|---|
| Comments | Osteostracans are known from the Silurian and Devonian of Europe, Siberia and North America. They are characterized by peculiar "cephalic fields" of unknown function, on the dorsal surface of the head-shield (red). Although jawless, they share with jawed vertebrates well-developed paired fins, an epicercal tail, cellular bone, and a sclerotic ring in eyes. Their mouth and gill opening are ventrally placed, as in galeaspids and pituriaspids. Their median, dorsal, nasohypophysial aperture, anterior to the eyes, is strikingly similar to that of lampreys but is now regarded as a convergence. All the osteostracans reconstructed here belong to the major clade Cornuata, whose generalized morphology is exemplified by the zenaspidid Zenaspis (bottom left). Some highly derived head-shield morphologies are exemplified by the benneviaspidids Hoelaspis (top right) and Tauraspis (top left), or the thyestiid Tremataspis (bottom right). The latter has lost the paired fins, possibly as a consequence of an adaptation to burrowing habits. |
| Reference | based on Janvier, P. 1985. Les Céphalaspides du Spitsberg. Anatomie, phylogénie et systématique des Ostéostracés siluro-dévoniens. Révision des Ostéostracés de la Formation de Wood Bay (Dévonien inférieur du Spitsberg). Cahiers de Paléontologie, Centre national de la Recherche scientifique, Paris. AND Janvier, P. 1985. Les Thyestidiens (Osteostraci) du Silurien de Saaremaa (Estonie). Première partie: Morphologie et anatomie, Annales de Paléontologie, 71(2), p.83-147. Deuxième partie: Analyse phylogénétique, répartition stratigraphique, remarques sur les genres Auchenaspis, Timanaspis, Tyriaspis, Didymaspis, Sclerodus et Tannuaspis. Annales de Paléontologie, 71(3), p.187-216. AND Mark-Kurik, E. & Janvier, P. 1995. Early Devonian osteostracans from Severnaya Zemlya, Russia. Journal of Vertebrate Paleontology. 15(3):449-462. |
| Image Use |
 This media file is licensed under the Creative Commons Attribution License - Version 3.0. This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 1997

|
About This Page
Muséum National d'Histoire Naturelle Paris, France
Page copyright © 1997
 Page: Tree of Life
Osteostraci.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
Osteostraci.
Authored by
Philippe Janvier.
The TEXT of this page is licensed under the
Creative Commons Attribution License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Citing this page:
Janvier, Philippe. 1997. Osteostraci. Version 01 January 1997 (under construction). http://tolweb.org/Osteostraci/14842/1997.01.01 in The Tree of Life Web Project, http://tolweb.org/

