




core Notholaena
Cloakferns




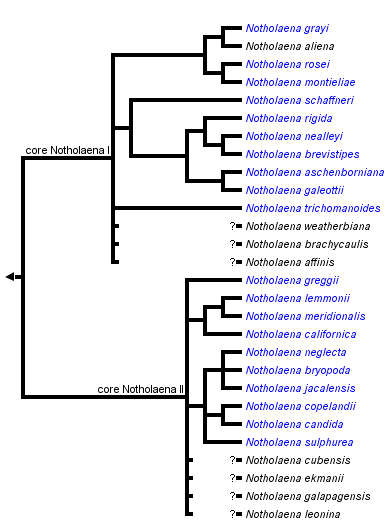


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Core Notholaena are comprised of approximately 27 species of small, farinose ferns (see below) of New World xeric habitats, with their center of richness in Mexico. As such, it contains the vast majority of Notholaena sensu Windham (1993a) and Mickel and Smith (2004) and Chrysochosma sensu Pichi Sermolli (1989). The sole known exception is Notholaena standleyi, which falls outside the core Notholaena clade based on recent DNA evidence (Rothfels et al., 2008). This circumscription also differs substantially from that of Tryon (1956)—many of the taxa included in his treatment are now known to be pellaeids, myriopterids, or hemionitids. For a more complete list of taxa previously included in Notholaena, see notholaenids: Excluded Species.
The genus Notholaena suffers from some unfortunate nomenclatural confusion: the genus was lectotypified three different times, and two of those lectotypifications (N. trichomanoides and N. marantae) are still in active use. Unfortunately, N. trichomanoides and N. marantae are only distantly related (N. trichomanoides is here, in core Notholaena, whereas N. marantae is in the pellaeids), and so the choice of one lectotype over the other results in a dramatically different concept of the genus. I here follow the first lectotypification (N. trichomanoides), based primarily on the arguments presented by Yatskievych and Smith (2003) and Rothfels et al. (2008), and treat N. marantae under Paragymnopteris.
Characteristics


Left: Notholaena californica ssp. leucophylla. Note the farina-covered leaf under-surfaces exposed when the leaves roll inwards under water stress. Note also the black marginal sporangia, and the absence of a false indusium. Right: Notholaena aschenborniana. The farina characteristic of all members of the core Notholaena are present in this species, too, but hidden under a dense layer of scales. Images © 2008 Carl Rothfels
This clade contains nearly half of all the world’s known farinose fern species. This farina can be brightly colored (usually white, but also cream, yellow, or orange) and is believed to be an adaptation to help reduce water loss from the leaves. Indeed, when these plants are water stressed, they roll their leaves inwards, exposing the farina-covered undersides to solar irradiation, even though the leaf undersurfaces are where the stomata are located (Hevly, 1963). All members of core Notholaena are farinose, even those, like N. aschenborniana, that have their farina covered by a dense layer of hairs or scales (the exception that proves the rule are the disjunct Chiapas populations of N. rigida, which are apparently nonfarinose; Mickel and Smith, 2004). Outside of the notholaenids the presence of farina is unusual, and it is unknown outside of the Pteridaceae (see A Quick Guide to Farinose Ferns for a brief overview of the genera of farinose ferns).
From other farinose species, core Notholaena taxa can generally be recognized by the position of their sporangia (confined to vein-ends near the leaf margins) and by the absence of a recurved leaf margin protecting the sporangia (although a weakly differentiated margin is present in some species). Confusion is most likely with members of Argyrochosma (the False Cloakferns) and with Notholaena standleyi (Northern Desert Star Cloakfern). Notholaena standleyi has a unique leaf shape and dissection (but see N. sulphurea). The farinose species of Argyrochosma differ from the highly dissected Notholaena species (e.g., N. greggii) in the following characters: the sporangia extend somewhat along the veins (vs. confined to the vein tips); rhizome scales are uniformly tan or brown (vs. rhizome scales uniformly dark or bicolored with dark central stripe); and glabrous gametophytes (vs. farinose).
Discussion of Phylogenetic Relationships
Based on plastid DNA sequence data (Rothfels et al., 2008), core Notholaena species are divided into two well-supported clades: core Notholaena I and core Notholaena II. Core Notholaena I species tend to have linear-lanceolate leaves and are often hairy or scaly, whereas the species in core Notholaena II tend to have pentagonal leaves (or, at least, enlarged basioscopic pinnules on their basal pinnae), and are usually glabrous (Rothfels et al., 2008).
References
Brown, R. 1810. Prodromus Florae Novae Holandiae et Insulae Van-Diemen. Richard Taylor & Sons, London.
Copeland, E. B. 1947. Genera Filicum. Chronica Botanica Co., Waltham.
Gastony, G. J., and D. R. Rollo. 1995. Phylogeny and generic circumscriptions of cheilanthoid ferns (Pteridaceae: Cheilanthoideae) inferred from rbcL nucleotide sequences. American Fern Journal 85:341-360.
Gastony, G. J., and D. R. Rollo. 1998. Cheilanthoid ferns (Pteridaceae: Cheilanthoideae) in the southwestern United States and adjacent Mexico--A molecular phylogenetic reassessment of generic lines. Aliso 17:131-144.
Giauque, M. F. A. 1949. Wax glands and prothallia. American Fern Journal 39:33-35.
Hall, C. C. 1950. Notholaena copelandii, a newly recognized species of the Texano-Mexican region. American Fern Journal 40:178--187.
Hevly, R. H. 1963. Adaptations of cheilanthoid ferns to desert environments. Journal of the Arizona Academy of Science 2:164-175.
Mickel, J. T. 1979. The fern genus Cheilanthes in continental United States. Phytologia 41:431-437.
Mickel, J. T., and A. R. Smith. 2004. The Pteridophytes of Mexico. The New York Botanical Garden Press, New York.
Pichi Sermolli, R. E. G. 1989. Again on the typification of the generic name Notholaena R. Brown. Webbia 43:301-310.
Rothfels, C. J., M. D. Windham, A. L. Grusz, G. J. Gastony, and K. M. Pryer. 2008. Toward a monophyletic Notholaena (Pteridaceae): Resolving patterns of evolutionary convergence in xeric-adapted ferns Taxon 57:712-724.
Seigler, D. S., and E. Wollenweber. 1983. Chemical variation in Notholaena standleyi. American Journal of Botany 70:790-798.
Tryon, R. M. 1956. A revision of the American species of Notholaena. Contributions from the Gray Herbarium 179:1-106.
Tryon, R. M., and A. F. Tryon. 1973. Geography, spores, and evolutionary relations in the cheilanthoid ferns. Pages 145-153 in The phylogeny and classification of ferns. (A. C. Jermy, J. A. Crabbe, and B. A. Thomas, eds.). Academic Press, New York.
Tryon, R. M., and A. F. Tryon. 1982. Ferns and Allied Plants with Special Reference to Tropical America. Springer-Verlag, New York.
Tryon, R. M., A. F. Tryon, and K. U. Kramer. 1990. Pteridaceae. Pages 404 in The Families and Genera of Vascular Plants (K. U. Kramer, and P. S. Green, eds.). Springer-Verlag, Berlin.
Windham, M. D. 1987. Argyrochosma, a new genus of cheilanthoid ferns. American Fern Journal 77:37-41.
Windham, M. D. 1993a. Notholaena. Pages 143--149 in Flora of North America (Flora of North American Editorial Committee, ed.) Oxford University Press, New York.
Windham, M. D. 1993b. Pteridaceae. Pages 122--124 in Flora of North America (Flora of North American Editorial Committee, ed.) Oxford University Press, New York.
Windham, M. D., L. Huiet, E. Schuettpelz, C. J. Rothfels, J. Beck, A. L. Grusz, G. Yatskievych, and K. M. Pryer. 2008. Using plastid and nuclear DNA sequences to redraw generic boundaries and demystify species complexes in cheilanthoid ferns. American Fern Journal (in review).
Windham, M. D., and G. Yatskievych. 2003. Chromosome studies of cheilanthoid ferns (Pteridaceae: Cheilanthoideae) from the western United States and Mexico. American Journal of Botany 90:1788-1800.
Wollenweber, E. 1984. Exudate flavonoids of Mexican ferns as chemotaxonomic markers. Rev. Latinoamer. Quim. 15:3-11.
Wollenweber, E., and H. Schneider. 2000. Lipophilic exudates of Pteridaceae -- chemistry and chemotaxonomy. Biochemical Systematics and Ecology 28:751-777.
Wollenweber, E., M. Dörr, and J. F. Stevens. 2001. A dihydroflavonol with taxonomic significance from the fern Notholaena sulphurea. Zeitschrift für Naturforschung 56:499-502.
Yatskievych, G., and A. L. A. Arbeláez. 2008. A new species and three generic transfers in the fern genus Notholaena (Pteridaceae). Novon 18:120-124.
Yatskievych, G., and A. R. Smith. 2003. Typification of Notholaena R. Br. (Pteridaceae). Taxon 52:331-336.
Title Illustrations

| Scientific Name | Notholaena nealleyi |
|---|---|
| Location | USA, Texas |
| Specimen Condition | Live Specimen |
| Identified By | Michael Windham |
| Image Use |
 This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0. This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2008 Carl Rothfels

|

| Scientific Name | Notholaena californica ssp. californica |
|---|---|
| Location | USA, Arizona, Black Canyon |
| Acknowledgements | Reproduced, with permission, from Arizona Ferns; azferns.org. |
| Specimen Condition | Live Specimen |
| Identified By | J.Metzgar |
| Life Cycle Stage | sporophyte |
| Image Use |
This media file is licensed under the Creative Commons Attribution License - Version 3.0.
|
| Copyright |
© 2005 Jordan Metzgar
|

| Scientific Name | Notholaena galeottii |
|---|---|
| Location | Queretaro, Mexico |
| Specimen Condition | Live Specimen |
| Identified By | C.J.Rothfels |
| Image Use |
This media file is licensed under the Creative Commons Attribution-NonCommercial License - Version 3.0.
|
| Copyright |
© 2009 Carl Rothfels
|
About This Page
Page copyright © 2008
 Page: Tree of Life
core Notholaena. Cloakferns.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
Page: Tree of Life
core Notholaena. Cloakferns.
The TEXT of this page is licensed under the
Creative Commons Attribution-NonCommercial License - Version 3.0. Note that images and other media
featured on this page are each governed by their own license, and they may or may not be available
for reuse. Click on an image or a media link to access the media data window, which provides the
relevant licensing information. For the general terms and conditions of ToL material reuse and
redistribution, please see the Tree of Life Copyright
Policies.
- First online 23 December 2008
- Content changed 07 January 2010
Citing this page:
Tree of Life Web Project. 2010. core Notholaena. Cloakferns. Version 07 January 2010. http://tolweb.org/core_Notholaena/133582/2010.01.07 in The Tree of Life Web Project, http://tolweb.org/

