




Diapriidae
Matthew Yoder


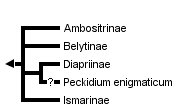


This tree diagram shows the relationships between several groups of organisms.
The root of the current tree connects the organisms featured in this tree to their containing group and the rest of the Tree of Life. The basal branching point in the tree represents the ancestor of the other groups in the tree. This ancestor diversified over time into several descendent subgroups, which are represented as internal nodes and terminal taxa to the right.

You can click on the root to travel down the Tree of Life all the way to the root of all Life, and you can click on the names of descendent subgroups to travel up the Tree of Life all the way to individual species.
For more information on ToL tree formatting, please see Interpreting the Tree or Classification. To learn more about phylogenetic trees, please visit our Phylogenetic Biology pages.
close boxIntroduction
Diapriids are parasitic wasps. They are abundant, diverse, and found world-wide (Masner, 1993). Most remain unnoticed to all but specialist collectors due to their small size, which averages ca. 2-3 mm in length. They are primarily solitary or gregarious endoparasitoids of Diptera (fly) larvae and pupae, though life histories for most species are unknown (Masner 1993, Chambers 1971). Some species are also known to attack ants (Loiacono 1987, Lachaud and Passera 1982) and staphylinid beetles (Masner 1993).
Immature stages are completely undescribed for nearly all species (but see Coon 2000, Menezes et al. 1998, Silvestri 1913). Several larger taxonomic treatises have been produced on the family (Masner and García 2002, Naumann 1980, Nixon 1980, Nixon 1957, Kieffer 1916), but to date less than half of the species estimated to occur worldwide (~4000) have been described (Masner 1993). Johnson's (1992) taxonomic catalog (see Information on the Internet) is an indispensable starting point for those studying diapriid systematics.
Characteristics
While diapriids have been treated as a natural group within the Apocrita there are presently few solid synapomorphies shared by all currently included members. Most characters used in identifying diapriids (see below) are likely plesiomorphic for the family. This absence of clear apomorphies is reflected in most family level keys (e.g. Masner 1993, Naumann and Masner 1985), which tend to key Diapriidae based on the absence of characters defining other potentially related families. The following characters, extended and modified from Masner (1993), are of use in distinguishing diapriids from the remaining Apocrita:
- Antennae inserted on a frontal prominence, or shelf that is not immediately next to the clypeus (except Ismarinae)
- First antennomere (scape) elongate, generally ~2x longer than wide
- Forewing without a clearly defined pterostigma
- Forewing with at most three closed cells
- Hindwing with at most one closed cell
- Males with antennomeres 3, 4, and 5, or combination thereof, with realease and spread structure (RSS, see Isidoro et al. 1999 for definition)
- Males and Females with fewer than 16 antennal segments, usually females with 12, 13 or 15 and males with 13 or 14
- Petiole with junction between sternite and tergite completely obliterated, generally tubular
Diapriids are currently classified into four subfamilies: Ismarinae, Ambositrinae, Belytinae and Diapriinae. The Ismarinae and Ambositrinae, most recently revised by Masner (1976) and Naumann (1982) respectively, are clearly defined morphologically and easily distinguished from all other diapriids. Ambositrines have metasomal tergites that are sharply reflexed laterally, and the second metasomal sternite (see anteroventral gaster in Fig. 1) is smaller than the third (Masner 1961). Ismarines have antennae that insert low on the face near to the clypeus and have a second, "false", spur on the protibia (Masner 1976).

Fig. 1. Representatives of the four diapriid subfamilies. Note: not all parts to scale. From top to bottom: Ismarinae- Ismarus sp.; Ambositrinae- Diphoropria sp.; Belytinae- Belyta sp.; Diapriinae- Idiotypa sp.
Antennal formulae (number of antennomeres, female-male) are 15-14 in Belytinae, Ambositrinae and Ismarinae, 12-14 or 13-13 in most Diapriinae. These differences can be used to separate most belytines from diapriines once Ismarinae and Ambositrinae are excluded. There are rarely a reduced numbers of segments in all subfamilies but the Ismarinae. Modified antennal segments in males (A3: Belytinae; A4 and/or A5: all others) further separate belytines from most, but not all, other diapriids. The Diapriinae are currently not defined by any apomorphy but rather as a group that is clearly not any of the other three.
Diapriids are nearly always clearly (i.e. characters of external morphology) sexually dimorphic, with the differences most notable in characteristics of the antennae. In this regard morpho-species studies involving diapriids must be particularly careful not to count conspecific males and females as separate species.
Discussion of Phylogenetic Relationships
Knowledge pertaining to the phylogenetic relationships of the family Diapriidae is limited. This is for the most part due to the relative paucity of research on the subject, rather than difficulties inherent to studying diapriids. The sister to Diapriidae is likely either the recently described Maamingidae (Early et al. 2001), Monomachidae, or Austroniidae (see Apocrita tree and references), though potential relationships with other apocritan superfamilies such as the Cynipoidea have been explored (Ronquist 1999). Phylogenetic analyses have not yet included the Ismarinae, who posses a unique morphology relative to the rest of the family. Their inclusion in future analyses should help resolve questions regarding both their relationship to the remaining diapriids and the relationships of diapriids to other apocritans.
Among the diapriid subfamilies the Belytinae is hypothesized as most basal (Masner, 1993), however its relationship to the Ismarinae and Ambositrinae is unclear (Masner 1961). It is highly probable that the Ismarinae and Ambositrinae are monophyletic while the boundary between the Belytinae and Diapriinae is less well-defined, being based primarily on characters associated with the loss or reduction of morphological features (Masner 1993, 1961).
The presence of a "belytine line" (a structure located on the metasomal sternites represented as a furrow or costa that limits the overlap of the ipsilateral metasomal tergites) has been suggested as a synapomorphy for the belytines (Masner 1993), however this feature is present in some Ambositrines and Ismarines (visible in Fig. 1) , and likely represents a plesiomorphy for Diapriidae or some larger containing clade.
Phylogenetic analyses treating taxonomic groups below the rank of family are currently limited to brief hypotheses for the subfamily Ambositrinae and to several other species group treatments (e.g. Loiacono and Maragaria 2000, Macek 1990, 1995, Masner 1991, Nauman 1982).
Diapriids currently contain several tenuously placed taxa such as Peckidium enigmaticum Masner and Garcia (2002). This taxon, while most likely a diapriid, is so apomorphic that some additional evidence, potentially molecular, will be needed to ensure its placement within the family. Also, to the untrained observer, several undescribed symphilic species are not easily placed within the Diapriidae, but these taxa are likely just highly apomorphic.
References
CHAMBERS, V.H. 1971. Large populations of Belytinae (Hymenoptera, Diapriidae). Entomologist's Monthly Magazine, 106: 149-154.
COON, B.R. 2000. Biology of Trichopria columbiana (Hymenoptera: Diapriidae), an endoparasitoid of Hydrellia pakistanae (Diptera: Ephydridae) a biological control agent of the aquatic weed Hydrilla verticillata (Hydrocharitaceae). Masters thesis. Entomology and Nematology Department, University of Florida, Gainesville.
EARLY, J.W., L. MASNER, A.D. AUSTIN AND I.I.D. NAUMANN. 2001. Maamingidae, a new family of proctotrupoid wasp (Insecta: Hymenoptera) from New Zealand. Invertebrate Taxonomy, 15: 341-352.
HOFFMEISTER, T. 1989. Biologie und Wirtskreis parasitischer Hautflügler der familie Diapriidae. Natur und Museum, 119: 327-334.
ISIDORO, N., F. BIN, R. ROMANI, J. PUJADE-VILLAR AND P. ROS-FARRE. 1999. Diversity and function of male antennal glands in Cynipoidea (Hymenoptera). Zoologica Scripta, 28: 165-174.
JOHNSON, N. 1992. Catalog of world Proctotrupoidea excluding Platygastridae. Memoirs of the American Entomological Institute, 51: 1-825.
KIEFFER, J.J. 1916. Diapriidae. Das Tierreich. Vol. 44.. Walter de Gruyter & Co., Berlin, 627 pp.
LACHAUD, J.P. AND L. PASSERA. 1982. Donnees sur la biologie de trois Diapriidae myrmecophiles: Plagiopria passerai Masner Solenopsia imitatrix Wasmann et Lepidopria pedestris Kieffer. Insectes Sociaux, 29: 561-568.
LOIACONO, M. S., 1987. Un nuevo diaprido (Hymenoptera) parasitoide de larvas de Acromyrmex ambiguus (Emery) (Hymenoptera, Formicidae) en el Uruguay. Revista de la Sociedad Entomologica Argentina, 44: 129-136.
MACEK, J. 1990. Revision of European Psilommina (Hymenoptera, Diapriidae). 1. Psilomma and Acanosema complex. Acta Entomologica Musei Nationalis Pragae [=Sborník Entomologického oddeleni N rodního Musea v Praze], 43: 335-360.
MACEK, J. 1995. Revision of the European species of Belyta Jurine. Acta musei Nationalis Pragae, Series B, Historia Naturalis, 51: 1-22.
MASNER, L. 1961. Ambositrinae, a new subfamily of Diapriidae from Madagascar and central Africa (Hymenoptera Proctotrupoidea). Memoires de l'Institut Scientifique de Madagascar, 12: 289-295.
MASNER, L. 1976. A revision of the Ismarinae of the New World (Hymenoptera, Proctotrupoidea, Diapriidae). Canadian Entomologist, 108: 1243-1266.
MASNER, L. 1991. Revision of Spilomicrus Westwood in America north of Mexico (Hymenoptera: Proctotrupoidea, Diapriidae). Canadian Entomologist, 123: 107-177.
MASNER, L. 1993. Superfamily Proctotrupoidea. pp.537-557 In JT Huber H Goulet (Eds),Hymenoptera of the world: An identification guide to families, Agriculture Canada, Ottawa, 668 pp.
MASNER, L. AND J.L. GARCÍA. 2002. The genera of Diapriinae (Hymenoptera: Diapriidae) in the New World. Bulletin of the American Museum of Natural History, 268: 1-138.
NAUMANN, I.D. 1982. Systematics of the Australian Ambositrinae (Hymenoptera: Diapriidae), with a synopsis of the non-Australian genera of the subfamily. Australian Journal of Zoology Supplementary Series No. 85: 1-239.
NAUMANN, I.D. AND L. MASNER. 1985. Parasitic wasps of the proctotrupoid complex: a new family from Australia and a key to world families (Hymenoptera: Proctotrupoidea sensu lato). Australian Journal of Zoology, 33: 761-783.
NIXON, G.E.J. 1957. Hymenoptera, Proctotrupoidea, Diapriidae, subfamily Belytinae. Handbooks for the Identification of British Insects no. 8(3dii): 1-107.
NIXON, G.E.J. 1980. Diapriidae (Diapriinae). Hymenoptera, Proctotrupoidea. Handbooks for the identification of British Insects no 8(3di): 1-55.
RONQUIST, F. 1999. Phylogeny, classification and evolution of the Cynipoidea. Zoologica Scripta, 28: 139-164.
SILVESTRI, F. 1913. Viaggio in Africa per cercare parassiti di mosche dei frutti. Bollettino del Laboratorio de Zoologia Generale e Agraria dell Facoltà Agraria in Portici, 8: 1-164.
Information on the Internet
Access to the electronic versions, including ongoing updates, of Johnson (1992) can be found at each of these links:
Hymenoptera Name Server at Ohio State.
Hymenoptera On-Line Database at Ohio State.
Title Illustrations

| Scientific Name | Spilomicrus sp. |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | M. Yoder |
| Sex | Male |
| Size | ~ 2mm |
| Collection | TAMUIC |
| Copyright |
© 2005 Matthew Yoder

|

| Scientific Name | Austroxylabis pictipennis |
|---|---|
| Specimen Condition | Dead Specimen |
| Identified By | M. Yoder |
| Sex | Female |
| Collection | TAMUIC |
| Copyright |
© 2005 Matthew Yoder
|
About This Page
Matthew Yoder
Texas A&M University, College Station, TX, USA
Correspondence regarding this page should be directed to Matthew Yoder at
Page copyright © 2005 Matthew Yoder
All Rights Reserved.
- First online 08 December 2004
- Content changed 22 February 2004
Citing this page:
Yoder, Matthew. 2004. Diapriidae. Version 22 February 2004. http://tolweb.org/Diapriidae/11312/2004.02.22 in The Tree of Life Web Project, http://tolweb.org/

